

| Journal of Clinical Gynecology and Obstetrics, ISSN 1927-1271 print, 1927-128X online, Open Access |
| Article copyright, the authors; Journal compilation copyright, J Clin Gynecol Obstet and Elmer Press Inc |
| Journal website http://www.jcgo.org |
Original Article
Volume 1, Number 4-5, October 2012, pages 63-66
The Effects of Ascorbic Acid on the Estrogen/Progesteron Levels in the Isolated Rabbit Uterine Muscle
Mehmet Suhha Bostancia, d, Merih Bayramb, Zuhal Aktuna Keskilc
aDepartment of Obstetrics and Gynecology, Sakarya Education and Research Hospital, Sakarya, Turkey
bDepartment of Obstetrics and Gynecology, Faculty of Medicine, Gazi University, Ankara, Turkey
cDepartment of Pharmacology, Faculty of Medicine, Kirikkale University, Kirikkale,Turkey
dCorresponding author: Mehmet Suhha Bostanci, Sakarya Egitim ve Arastırma Hastanesi Kadin Hastaliklari ve Dogum Klinigi Sakarya Turkey
Manuscript accepted for publication October 4, 2012
Short title: Effects of Ascorbic Acid
doi: https://doi.org/10.4021/jcgo47w
| Abstract | ▴Top |
Background: The study was carried out to examine the effect of Ascorbic Acid (AsA) on estrogen/progesterone levels in the isolated non-pregnant rabbit uterine muscle.
Methods: Twelve non-pregnant rabbits were randomized into two groups as the rabbits pretreated with AsA (n = 6) and the rabbits not pretreated with AsA (n = 6). After 30 mg/kg AsA was administered intraperitoneally, serum and tissue (uterine smooth muscle) levels of estrogen and progesterone were measured.
Results: AsA at the above mentioned concentration caused a significant increase in tissue estrogen/progesterone ratio (P < 0.01), while did not induce any change in the hormone ratio in the serum. The increase in the tissue estrogen/progesterone ratio was resulted from both the decrease of the progesterone levels and the increase of estrogen levels (P < 0.001).
Conclusion: In this context, the fact that the tissue levels of hormones and autacoids is more important than the serum concentrations should be concidered. The findings of this study indicate that in comparison to the control group, AsA causes decrease in the non-pregnant isolated uterine smooth muscle tissue levels of progesterone and increase in the non-pregnant isolated uterine smooth muscle tissue levels of estrogen while the serum levels keep constant.
Keywords: Ascorbic acid; Uterine muscle; Estrogen; Progesterone
| Introductıon | ▴Top |
Estrogen and progesterone are steroid sex hormones which are important for the maintenance of uterine quiescence, for the stimulation of myometrial gap junction formation, for the regulation of uterine blood flow and the synthesis of cell-surface reseptors [1]. An alleged risk of long-term estrogen use is the increased incidence of breast cancer [1].
Estrogen influence on target tissues is the most important problems in understanding of the estrogen-induced carsinogenesis and anticarsinogenesis mechanisms. Some conditions and factors (Ascorbic acid, alpha-tocopherol, etc.) increasing the genotoxic component in total effect of estrogens on the uterine tissue are very important, since these factors may influence both the hormonal carsinogenesis type and biological properties of the developing hormone-dependent tumors [2] Alpha-tocopherol and ascorbic acid (AsA) have been suggested to play a role in breast cancer prevention due to their antioxidative capacity. Increased exposure to endogenous and exogenous sex steroids is a known risk factor for breast cancer [3]. Ascorbate inhibits oxygen consumption when estradiol is metabolized in the presence of either glutathione or nicotinamide adenine dinucleotide (NADH) by reducing radical intermediates back to their parent molecules and forming the relatively stable ascorbate radical. The accumulation of intracellular hydrogen peroxide could explain the hydroxyl radical-induced DNA base lesions recently reported for female breast cancer tissue [4].
At the cellular level, theoretical evidence supports the idea that estrogen itself may serve as an antioxidant, although much of the work examines estrogen's ability to protect membrane integrity rather than oxidative stress directly. It appears that women taking oral contraceptives exhibited higher erythrocyte glutathione peroxidase activity [5]. This finding suggests that differences in endogenous plasma estrogen may affect lipid peroxidation and antioxidant enzyme status because exogenous steroids are thought to be associated with increased free radicals, lipids, and lipid peroxides in the blood [6]. This theory was contradicted by Kanaley and Ji, who found that erythrocyte glutathione peroxidase activity was significantly higher in amenorrheic athletes relative to eumenorrheic control subjects [6]. The authors suggest, however, that the observed differences in glutathione peroxidase activity may not be a direct result of estrogen status, but rather estrogen status may influence blood antioxidant activity through another mechanism, such as plasma iron concentrations [7]. In any case, it appears that estrogen status and blood antioxidant capacity are related in some way.
Vitamin C is water soluble and can directly react with superoxide, hydroxyl radicals, and singlet oxygen [8]. Byrd et al have suggested AsA to be associated with steroidogenesis in chickens as have been indicated for mammalian species. They have reported that AsA had caused an increase in progesterone biosynthesis by activating luteinizing hormone [9]. AsA does have an important role as a modulator of aromatase activity in vivo in rats [10].
The present study was undertaken to examine the effects of AsA on estrogen and progesterone levels in the isolated non-pregnant rabbit uterine muscle and serum, in order to assess the modulatory effect of AsA on hormone levels that may increase cost-effectiveness and decrease adverse effects of hormonal therapies especially postmenopausal therapy.
| Materıals and Methods | ▴Top |
This is the experimental study that has been made at the University of Gazi Medical School, Department of Pharmacology Research Laboratory and Kirikkale University Medical School, Department of Gynecology and Obstetrics.
Study groups
Twelve non-pregnant adult, New Zeland female rabbits weighing 3.0 - 3.5 kg were kept in standard laboratory conditions; at 25 °C under controlled lighting (12 hours light, 12 hours darkness). They were given a standard diet and tap water ad libitum. In terms of Helsinki declaration, standart care was taken for all the animals. The experiments complied fully with the current laws of our country.
Group 1 (n = 6): The rabbits were pretreated with AsA, 30 mg/kg body weight intraperitoneally for five days. (This application way was choosen because of preventing the insufficiency absorbtion of ascorbic acid from digestive tract).
Group 2 (n = 6): As the control rabbits, they were injected the same volume of saline intraperitoneally for five days.
Surgical procedures
The animals were anaesthetised with sodium pentobarbital (30 mg/kg I.V.) and an abdominal midline incision was performed to all groups’ animals, and then the uterus were quickly removed, the endometrium was gently dissected from myometrium and isolated uterine smooth muscle was transferred to a tube for extraction. Blood samples were taken directly from the heart and transferred to the tubes for centrifugation at 3,000 g.
Laboratory assay
Tissue specimens in isotonic solution were homogenized by doing 8 stroke/min with teflon capsulated homogenizatour, and then they were centrifugated. Estrogen and progesterone assay were estimated from supernatant.
Estrogen and progesterone levels in both tissue and serum were estimated by using RIA CT Radium Radioimmunuasay for estrojen and Immulite 2000 Chemiluminescent technique for progesterone. The results were given as pg/mL for estrogens and ng/mL for progesterone at serum. In wet weight of tissue, the results were given as pg/g for estrogen and ng/g for progesterone.
Statistical analysis
All the results are expressed as mean ± SEM. The values of differences between groups were compared using the Student’s t-test. Values were considered significant at P < 0.05.
| Results | ▴Top |
Tissue Progesterone levels of group 1 was found to be 0.0005 ± 0.00008 ng/g (wet weight) while it was 0.01 ± 0.002 ng/g for group 2. The lower level of the tissue progesterone in the AsA pretreated group was found to be statistically significant (P < 0.001). Progesterone levels in the AsA pretreated and the control group were 1.4 ± 0.1 ng/mL and 1.5 ± 0.4 ng/mL for the serum respectively. No statistical difference existed in terms of serum progesterone levels due to AsA treatment. Estrogen levels both in the tissue and serum were compared. Tissue estrogen levels were found to be 0.13 ± 0.03 pg/g (wet weight) for group 1 and 0.08 ± 0.01 pg/g for group 2 and in the AsA pretreated group this situation was significantly higher than the control group (P < 0.05). Serum estrogen levels for group 1 and group 2 were 101.7 ± 3.1 pg/mL and 98.1 ± 9.8 pg/mL, respectively. There was no statically difference between two groups for serum estrogen levels (Table 1).
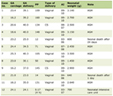 Click to view | Table 1. Tissue (Wet Weight) and Serum Progesteron and Estrogen Levels of the Groups |
Tissue estrogen/progesterone ratio was found to be 188.9 ± 68.4 in the group 1 and 8.45 ± 3.15 in the group 2. Higher tissue estrogen/progesterone ratio in the AsA pretreated group was found to be statistically significant (P < 0.001). Serum estrogen/progesterone ratio was 80.3 ± 7.6 for AsA pretreated group and 76.3 ± 4.4 for the control group which is found to be statistically insignificant (Table 2).
Click to view | Table 2. Tissue (Wet Weight) and Serum Estrogen and Progesteron Level Ratios of the Groups |
| Discussion | ▴Top |
According to the results of the present study, AsA caused a significant decrease in tissue progesteron levels while did not change serum progesterone concentrations in the non-pregnant rabbits. While increased tissue levels of estrogen, serum concentrations of estrogen did not change by AsA pretreatment.Although AsA has been shown to have an important role as a modulator of aromatase activity in rats in-vivo and altered the binding and enzymatic activities of estrogen [10]. AsA caused a significant decrease in tissue progesterone levels while did not change serum progesterone concentrations. The decrease in tissue progesterone level might depend to the activation of prostaglandin synthesis via cyclooxygenase by AsA [11]. AsA has been reported to inhibit prostaglandin synthesis by removing intermediate radicals that are essential for cyclooxygenase mechanisms [12]. It has been shown that prostaglandin F2α produced a decrease in progesterone levels [13]. However, serum progesterone concentrations did not change by AsA. Therefore the decrease in tissue progesterone levels could not be explained by the activation of prostaglandin synthesis by AsA. These results indicate that vitamin C may alter tissue levels of progesterone prior to its effect on circulating levels and can be assumed that AsA promotes the degradation of progesterone via increasing activity of the enzymes involved in the tissue. Since it is well known that AsA is involved in steroid biosynthesis.
Combinations of vitamins C and E enhances some features of estrogen effects (uterine weight, induction of progesterone receptors) but attenuates the other (proliferation index). So that the combination of N-acethylcysteine and optimal doses of ascorbic scid and alpha-tocopherol may be recommended for prevention of the phenomenon of switching of estrogen effects [2]. The alteration of luteinising hormone (LH) by AsA might be due to the decreased activity of progesterone which caused a negative feedback inhibition on LH.
Nutritional deficiency of AsA increases by oral contraseptives with low-dose-estrogen [14]. But vitamin C, an important factor in uterine estrogen binding, does not have a role in the prevention of Depo-Provera-induced menstruel irregularities [15].
Ascorbic acid may interact with estrogen therapy that this effect of AsA with estradiol (E2) may be possibly antioxidant [16]. Studies in animals support the theory that estrogen protects membrane integrity during times of oxidative stress. Antioxidants act as a cooperative network, employing a series of redox reactions [17-20]. Additive effect of AsA in anti-oxidation may be responsible for increased levels of tissue estrogen. As a result of the study, ascorbic acid has been interacted with estrogen therapy, this effect may be found possible interaction of AsA with E2 at the level of antioxidation [16].
As a conclusion, the finding of this study showed that AsA in comparison to control group, while did not have an effect on serum estrogen/progesterone ratio, induced the increased tissue estrogen/progesterone ratio. This catalisor effect of AsA at tissue may help minimising hormone-induced morbidity by adjusting clinically used hormone dosage.
| References | ▴Top |
- Speroff L, Glass RH, Kase NG. Clinical Gynecologic Endocrinology and Infertility. Sixth Edition. Lippincott Williams & Wilkins,Philadelphia. 1999:1-3. Book Chapters.
pubmed - Berstein L, Tsyrlina E, Poroshina T, Bychkova N, Kalinina N, Gamajunova V, Vasilyev D, et al. Switching (overtargeting) of estrogen effects and its potential role in hormonal carcinogenesis. Neoplasma. 2002;49(1):21-25.
pubmed - Dabrosin C, Ollinger K. Protection by alpha-tocopherol but not ascorbic acid from hydrogen peroxide induced cell death in normal human breast epithelial cells in culture. Free Radic Res. 1998;29(3):227-234.
doi - Sipe HJ, Jr., Jordan SJ, Hanna PM, Mason RP. The metabolism of 17 beta-estradiol by lactoperoxidase: a possible source of oxidative stress in breast cancer. Carcinogenesis. 1994;15(11):2637-2643.
doi pubmed - Capel ID, Jenner M, Williams DC, Donaldson D, Nath A. The effect of prolonged oral contraceptive steroid use on erythrocyte glutathione peroxidase activity. J Steroid Biochem. 1981;14(8):729-732.
doi - Kanaley JA, Ji LL. Antioxidant enzyme activity during prolonged exercise in amenorrheic and eumenorrheic athletes. Metabolism. 1991;40(1):88-92.
doi - Jenkins RR, Krause K, Schofield LS. Influence of exercise on clearance of oxidant stress products and loosely bound iron. Med Sci Sports Exerc. 1993;25(2):213-217.
doi pubmed - Sauberlich HE. Ascorbic acid. In: Brown ML, ed. Present knowledge in nutrition. Washington, DC: International Life Sciences Institute.1993;132–141.
- Byrd JA, Pardue SL, Hargis BM. Effect of ascorbate on luteinizing hormone stimulated progesterone biosynthesis in chicken granulosa cells in vitro. Comp Biochem Physiol Comp Physiol. 1993;104(2):279-281.
doi - Densmore CL, Schauweker TH, Gregory RR, Webb B, Garcia E, Markaverich BM. Chromatographic resolution of the type II estrogen binding site and a tyrosinase-like enzymatic activity from rat uterine nuclei. Steroids. 1994;59(4):282-287.
doi - Samuelsson B. Biosynthesis of prostaglandins. Fed Proc. 1972;31(5):1442-1450.
pubmed - Honjo H. [Endogenous levels and dynamics of estrogen sulfates—physiological and pathological roles of estrone sulfate and estradiol 17-sulfate]. Nihon Naibunpi Gakkai Zasshi. 1992;68(11):1158-1166.
pubmed - Aten RF, Duarte KM, Behrman HR. Regulation of ovarian antioxidant vitamins, reduced glutathione, and lipid peroxidation by luteinizing hormone and prostaglandin F2 alpha. Biol Reprod. 1992;46(3):401-407.
doi pubmed - Murthy NK, Vijaya S. Vitamin nutritional status of women using oral contraceptive pills. Indian J Nutr Diet. 1980;17(3):79-83.
pubmed - Harel Z, Biro F, Kollar L, Riggs S, Flanagan P, Vaz R. Supplementation with vitamin C and/or vitamin B(6) in the prevention of Depo-Provera side effects in adolescents. J Pediatr Adolesc Gynecol. 2002;15(3):153-158.
doi - Vihtamaki T, Parantainen J, Koivisto AM, Metsa-Ketela T, Tuimala R. Oral ascorbic acid increases plasma oestradiol during postmenopausal hormone replacement therapy. Maturitas. 2002;42(2):129-135.
doi - Amelink GJ, Koot RW, Erich WB, Van Gijn J, Bar PR. Sex-linked variation in creatine kinase release, and its dependence on oestradiol, can be demonstrated in an in-vitro rat skeletal muscle preparation. Acta Physiol Scand. 1990;138(2):115-124.
doi pubmed - Amelink GJ, van der Wal WA, Wokke JH, van Asbeck BS, Bar PR. Exercise-induced muscle damage in the rat: the effect of vitamin E deficiency. Pflugers Arch. 1991;419(3-4):304-309.
doi pubmed - Bar PR, Amelink GJ, Oldenburg B, Blankenstein MA. Prevention of exercise-induced muscle membrane damage by oestradiol. Life Sci. 1988;42(26):2677-2681.
doi - Ayres S, Baer J, Subbiah MT. Exercised-induced increase in lipid peroxidation parameters in amenorrheic female athletes. Fertil Steril. 1998;69(1):73-77.
doi
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Clinical Gynecology and Obstetrics is published by Elmer Press Inc.